
你以為你看到了世界,其實你看到的是大腦的猜測
從轉導、拓樸地圖到預測編碼,看視聽觸味嗅五大感覺如何把物理能量建構成你的主觀世界。
你以為你看到了世界,其實你看到的是大腦的猜測
請你閉上一隻眼睛,把一根手指放在面前,慢慢往鼻尖移動。你會發現手指逐漸變得模糊、甚至「分裂」成兩根。但在日常生活中,你幾乎從不抱怨世界是「兩個」的。為什麼?因為你「看到」的世界,並不是眼睛拍下來的照片,而是大腦根據視網膜上殘缺、扭曲、晃動的訊號,即時「推論」出來的最佳猜測(best guess)。
這正是感覺系統(sensory systems)最迷人的地方。視(vision)、聽(audition)、觸(somatosensation)、味(gustation)、嗅(olfaction)這五大感覺,每一種都不是被動的「接收器」,而是一套主動建構現實的神經運算系統。它們把外在世界的物理能量——光波、聲波、壓力、化學分子——轉換成神經元能聽懂的語言(動作電位),再經過層層處理,最終讓你「感覺」到一杯咖啡的香氣、溫度與苦味。
這篇文章將帶你走過這趟「從物理世界到主觀經驗」的旅程。我們會看到五種感覺背後共享的設計原則,也會看到它們各自精巧的解法。
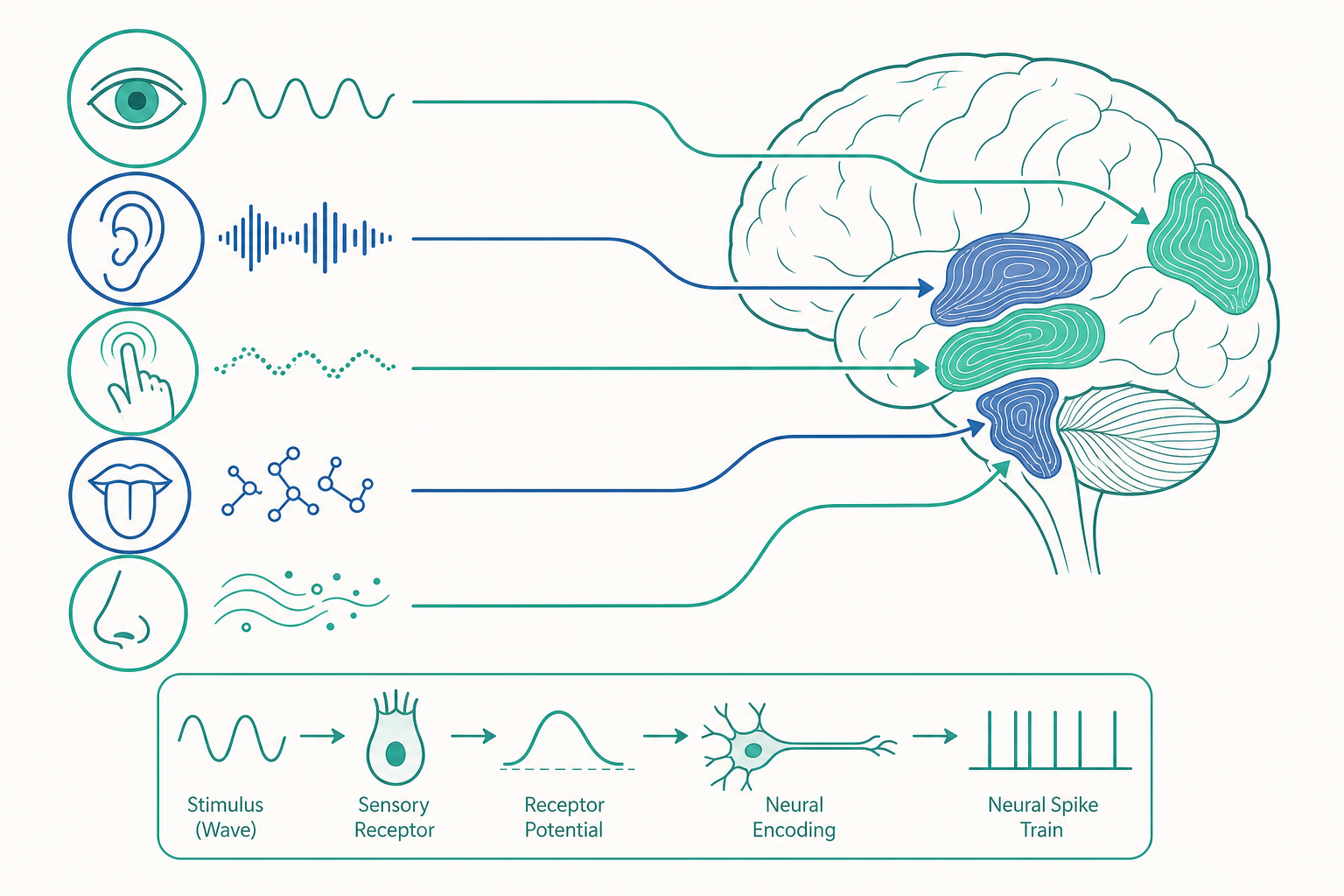
一切的起點:轉導(transduction)
不論哪一種感覺,第一步都是相同的問題:如何把外界的物理或化學能量,變成神經系統能處理的電訊號?這個過程叫做感覺轉導(sensory transduction),由特化的感受器細胞(receptor cells)負責。
每一種感覺有它專屬的轉導機制:
- 視覺:視網膜上的視桿細胞(rod)與視錐細胞(cone)含有光色素(photopigment,如視紫質 rhodopsin)。一個光子打中視紫質,會讓其中的視黃醛(retinal)發生形狀改變(11-順式變成全反式),觸發 G 蛋白偶聯的訊號級聯,最終改變細胞膜電位。
- 聽覺:耳蝸(cochlea)內的毛細胞(hair cell)頂端有一束纖毛(stereocilia)。聲波引起的振動使纖毛偏折,直接拉開機械門控離子通道(mechanically gated channel),讓鉀離子湧入。這是最快速的轉導之一——幾乎沒有第二信使的延遲。
- 觸覺:皮膚中的機械感受器(如 Merkel 細胞、Pacinian 小體)含有 Piezo2 通道,受壓力形變時開啟。發現 Piezo 通道的 Ardem Patapoutian 與發現溫度/辣感受器 TRPV1 的 David Julius 共同獲得 2021 年諾貝爾生理醫學獎。
- 味覺:舌頭味蕾中的味覺細胞,對甜、鮮、苦使用 G 蛋白偶聯受體(GPCR,如 T1R、T2R 家族),對鹹、酸則用離子通道直接偵測鈉離子與氫離子。
- 嗅覺:鼻腔嗅上皮的嗅覺神經元,每個只表現一種嗅覺受體(olfactory receptor)。Linda Buck 與 Richard Axel 因發現這個龐大的受體基因家族(人類約 400 個功能性基因)獲得 2004 年諾貝爾獎。
設計原則:適當刺激(adequate stimulus)。 每種感受器只對特定能量形式最敏感。但這也帶來一個哲學意涵——你感覺到的「光」或「聲音」,其實是大腦對「哪一條神經被活化」的詮釋。用手指按壓眼球會「看到」光點(眼內閃光,phosphene),正是因為視覺通路被機械刺激活化了,大腦仍然把它解讀成「光」。
第二個共同設計:拓樸地圖(topographic maps)
感覺資訊進入大腦後,並不是隨機散落,而是有秩序地排列成地圖。相鄰的感受器,通常投射到相鄰的腦區。
- 視覺:初級視覺皮質(V1)保留了視網膜的空間排列,稱為視網膜拓樸(retinotopy)。
- 聽覺:耳蝸從底端到頂端依序對不同頻率敏感(底端高頻、頂端低頻),這個頻率排列一路保留到聽覺皮質,稱為音調拓樸(tonotopy)。
- 觸覺:體感皮質上身體各部位依序排列,形成著名的皮質小人(cortical homunculus)。嘴唇、手指佔據的皮質面積遠大於背部或大腿——因為這些部位的感受器密度高、需要更精細的處理。
這些地圖不是按身體實際比例,而是按「感受器密度」與「行為重要性」分配資源。這是神經系統「把運算資源花在刀口上」的具體展現。
邊緣增強:側抑制(lateral inhibition)
感覺系統不只是忠實地傳遞訊號,它還會「加工」訊號來凸顯重要資訊。最經典的機制是側抑制(lateral inhibition):一個被活化的神經元,會抑制它旁邊的神經元。
這帶來什麼好處?強化邊界與對比。想想看你在閱讀這段文字時,黑字與白底之間的邊界看起來特別銳利——側抑制讓邊界兩側的明暗差異被放大了。
看一個例子:馬赫帶(Mach bands)
請你想像一組由淺到深、一階一階排列的灰色長條(灰階階梯)。盯著它看,你會發現每一個色塊的「邊緣」似乎不太均勻——靠近較暗色塊的那一側看起來更亮一點,靠近較亮色塊的那一側看起來更暗一點。這些並不存在於實際亮度中的亮帶與暗帶,就叫做馬赫帶(Mach bands),由物理學家 Ernst Mach 在 19 世紀描述。
馬赫帶是側抑制的直接後果。在亮度均勻的區域,神經元彼此抑制的程度相當,輸出穩定;但在邊界處,較亮側的神經元受到較暗鄰居的抑制較少(因此輸出被「拉高」),較暗側則受到較亮鄰居的強抑制(輸出被「壓低」)。結果就是邊界被誇大了。
這個例子告訴我們一件重要的事:你的知覺並不等於物理現實。感覺系統是在「忠實」與「有用」之間做權衡,而它選擇了「有用」——凸顯邊界,幫助你辨識物體輪廓。
從特徵到物件:階層式處理(hierarchical processing)
感覺皮質採用階層式(hierarchical)架構:低層處理簡單特徵,高層整合成複雜的物件表徵。
視覺系統是研究得最透徹的例子。David Hubel 與 Torsten Wiesel 在 1950–60 年代用微電極記錄貓的視覺皮質,發現 V1 的神經元並非對「點狀光」反應,而是對特定方向的線條或邊緣反應——這就是方向選擇性(orientation selectivity)。他們因此獲得 1981 年諾貝爾獎。
往高層走,神經元的偏好越來越複雜:
- V1:朝向特定角度的邊緣、線條。
- V2、V4:輪廓、簡單形狀、顏色。
- 下顳葉皮質(IT cortex):完整物件,甚至有對「臉」高度選擇性的神經元(如梭狀回臉孔區 FFA)。
這條路徑被稱為腹側流(ventral stream)或「what 路徑」,負責「這是什麼」。另一條背側流(dorsal stream)或「where/how 路徑」,則負責空間位置與動作引導。一位視覺失認症(visual agnosia)的著名病人 D.F. 能準確地把信投進不同角度的信箱(背側流完好),卻說不出信箱口是什麼方向(腹側流受損)——這個雙重解離(double dissociation)有力地支持了兩條路徑的分工。
嗅覺與味覺:化學感覺的特殊性
視聽觸三種感覺處理的是物理能量(光、聲、壓),而嗅覺與味覺處理的是化學分子,因此設計原則略有不同。
嗅覺沒有像視網膜那樣整齊的空間地圖,而是用組合編碼(combinatorial coding):一種氣味分子會活化多個受體,一個受體也會回應多種分子。大腦透過「哪些受體被活化、活化多強」的組合模式來辨識氣味。這讓人類雖然只有約 400 種嗅覺受體,卻能分辨上萬種(有研究估計甚至上兆種)氣味。嗅覺也是唯一不經視丘(thalamus)中繼就能直接抵達皮質與邊緣系統的感覺——這解釋了為什麼氣味能如此強烈、即時地勾起情緒與記憶(普魯斯特現象)。
味覺常被誤解。一個常見迷思是「舌頭分區地圖」——說舌尖嚐甜、兩側嚐酸、舌根嚐苦。這是 20 世紀初一篇德文論文被誤譯放大的結果,已被證實是錯的:舌頭上每個味蕾區域都能偵測全部五種基本味道(甜、鹹、酸、苦、鮮 umami),只是敏感度有些微差異。此外,我們平常說的「味道」(flavor)其實是味覺、嗅覺、口感與溫度的整合——所以感冒鼻塞時,食物會變得「沒味道」,正是因為嗅覺被堵住了。
主動推論:感覺不是單行道
最後一個關鍵概念,也是當代神經科學最重要的轉變之一:感覺處理不是由下而上(bottom-up)的單行道,而是雙向的。
從感受器往上傳的訊號固然重要,但從高層皮質往下傳的回饋連結(feedback projections)數量同樣龐大,甚至更多。大腦會根據過去經驗與當前脈絡,預測接下來會收到什麼感覺輸入,再拿實際輸入與預測比對。這個框架叫做預測編碼(predictive coding)或主動推論(active inference)。
這也是為什麼回到開頭那句話——你看到的世界是大腦的「最佳猜測」。當預測與輸入吻合,你看到穩定的世界;當兩者衝突,你就會經驗到錯覺、驚訝,甚至幻覺。
重點回顧
- 轉導(transduction) 是所有感覺的共同起點:特化的感受器把光、聲、壓力、化學分子等物理/化學能量,轉換成神經電訊號。每種感覺有專屬機制(光色素、機械門控通道、GPCR 等)。
- 拓樸地圖(topographic maps) 讓感覺資訊有秩序地排列(視網膜拓樸、音調拓樸、皮質小人),且資源按感受器密度與行為重要性分配,而非身體實際比例。
- 側抑制(lateral inhibition) 凸顯邊界與對比,馬赫帶錯覺就是它的直接證據——提醒我們知覺是「有用」而非「忠實」的建構。
- 階層式處理(hierarchical processing) 從簡單特徵(V1 的方向選擇性)逐層整合為複雜物件(IT 皮質的臉孔細胞),並分化為腹側「what」與背側「where/how」兩條流。
- 嗅味覺用化學編碼:嗅覺靠組合編碼分辨上萬種氣味且直達邊緣系統;味覺的「舌頭分區地圖」是迷思,flavor 是多感覺整合的結果。
深入探討(研究所視角)
1. 預測編碼的計算神經機制。 預測編碼(predictive coding)由 Rao 與 Ballard(1999)形式化,後由 Karl Friston 的自由能原理(free energy principle)統整為更宏大的理論框架。其核心主張是:感覺皮質的階層中,每一層都試圖預測下一層較低階的活動,只有預測誤差(prediction error)會往上傳。這在解剖學上可能對應到皮質的層狀結構——前饋(feedforward)連結多源自表淺層(傳遞預測誤差),回饋(feedback)連結多源自深層(傳遞預測)。一個可檢驗的推論是重複抑制(repetition suppression):可預測的刺激引發較弱的神經反應,因為預測誤差小。這已在 fMRI 與單細胞記錄中被觀察到。這個框架把感知、注意、學習統一在「最小化預測誤差」的單一目標下,與機器學習中的變分推論(variational inference)有深刻的數學對應。
2. 與 AI 的雙向啟發。 Hubel 與 Wiesel 的階層式視覺發現,直接啟發了 Fukushima 的 Neocognitron(1980)與後來的卷積神經網路(CNN)——「簡單特徵→複雜特徵」的逐層抽象,正是深度視覺網路的核心。反過來,近年研究(如 Yamins & DiCarlo 等人)發現,訓練得好的深度網路,其中間層的表徵竟能高度預測獼猴 IT 皮質的神經反應,使 CNN 成為視覺皮質的「計算模型」。這形成一個迷人的循環:神經科學啟發 AI,AI 又回頭成為理解大腦的工具。不過要謹慎——表徵相似不等於機制相同,生物視覺的回饋連結、時序動態與能耗效率,目前的前饋網路仍遠遠無法匹配。
3. 跨感覺整合與可塑性。 感覺系統並非彼此隔離。多感覺整合(multisensory integration)在上丘(superior colliculus)與聯合皮質發生,遵循「時空一致性」與「逆效律(inverse effectiveness)」——當單一感覺訊號越微弱,多感覺整合帶來的增益越大。麥格克效應(McGurk effect,看到的嘴型改變你聽到的音節)與腹語術錯覺都是證據。更戲劇性的是跨感覺可塑性(cross-modal plasticity):先天失明者的視覺皮質會被聽覺與觸覺「徵用」,這也是盲文閱讀與回聲定位的神經基礎。這對連結優心理學(知覺、注意理論)與優生物學(神經發育、關鍵期可塑性)提供了豐富的交會點:感覺地圖在發育關鍵期高度可塑,經驗會雕塑突觸連結(如 Hubel & Wiesel 的單眼剝奪實驗顯示視覺剝奪會永久改變眼優勢柱)。
延伸思考方向: 若你對「主觀經驗如何從神經活動湧現」感興趣,這正通往意識研究(consciousness studies)的核心難題——感覺處理告訴我們神經如何表徵世界,但為何這些表徵伴隨著「感覺起來像什麼」(qualia)的主觀經驗,至今仍是開放問題。從感覺系統出發,你已站在認知神經科學最深邃疑問的門口。